在微观生态中,细菌与噬菌体之间存在着持续的共同进化与博弈。为了抵御噬菌体的感染,细菌演化出了复杂的防御网络。除了CRISPR-Cas系统外,广泛存在于原核生物中的“流产感染”(Abortive infection, Abi)也是一种重要的防御策略。这是一种群体保护机制:当细菌检测到噬菌体感染时,会启动程序性细胞死亡或生长停滞,通过牺牲受感染的个体来阻断病毒的复制与扩散,从而保护菌群中的其他同伴。
Lamassu(Lmu)系统是2018年首次被鉴定出的一种Abi防御系统。该系统主要由效应蛋白(LmuA)和DNA传感器(LmuB)组成,部分II型变体还包含一个辅助组分LmuC。尽管此前研究已预测LmuB属于SMC家族,而LmuA具有核酸酶等结构域,但该系统具体的组装模式、感应机制以及激活过程一直未被完全阐明 。
2026年1月2日,中国科学技术大学张凯铭团队与北京化工大学冯越/张怡团队合作在Nature Chemical Biology上发表了题为Structural insights into type-I and type-II Lamassu antiphage systems的研究论文 。该研究解析了I型和II型Lamassu系统在多种状态下的高分辨率冷冻电镜结构,结合生化实验与功能验证,详细阐明了该系统的组装方式及其精细的调控机制(图1)。
研究团队首先明确了Lamassu系统的整体架构:其核心是由SMC家族蛋白LmuB构成的二聚体支架 。在II型系统中,LmuC位于复合物中心 。在未受感染的静息状态下,具有核酸酶活性的LmuA以单体形式被LmuB的螺旋结构域紧密结合,这种构象将LmuA锁定在非活性状态,避免了对细菌自身DNA的误伤 。
通过解析Lamassu系统与DNA底物的复合物结构,团队揭示了其独特的感应机制:与传统SMC复合物环绕DNA的方式不同,Lamassu系统通过特异性识别双链DNA的“末端”来感知噬菌体入侵(DNA末端通常在噬菌体复制或基因组受损时大量产生)。此外,研究还发现该系统存在一种“双重检查”机制:高浓度ATP(代表细胞代谢正常)会抑制核酸酶活性,而低浓度ATP(暗示噬菌体感染导致的能量耗竭)则会促进系统与DNA结合并切割DNA 。一旦被激活,噬菌体DNA的结合会导致LmuA从LmuB支架上释放。释放后的LmuA单体会在细胞质中迅速发生构象变化,组装成具有高活性的四聚体。该四聚体能够无差别地切割细胞内的DNA(包括宿主和噬菌体的DNA),最终导致受感染细菌死亡,从而阻断噬菌体的传播。
综上,本研究不仅展示了Lamassu系统从静息到激活的完整分子图像,还揭示了SMC家族蛋白在原核生物免疫中的全新功能——不仅是染色体的维护者,更是免疫系统的核心支架。通过解析LmuA从单体到四聚体的激活路径,为理解细菌如何利用多聚化机制控制免疫应答提供了新的理论框架。
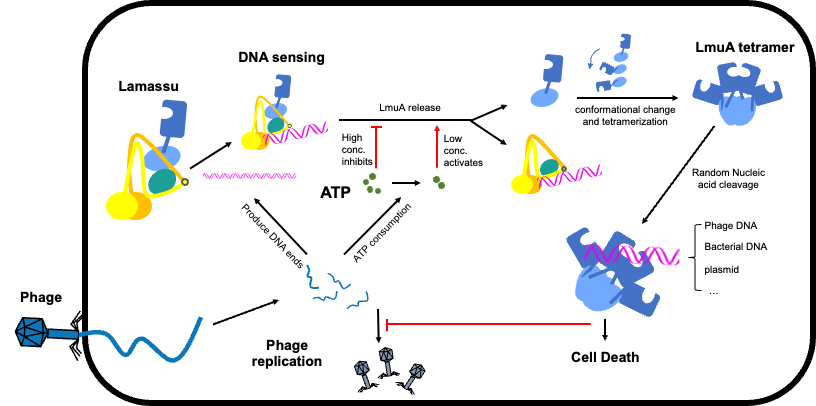
图1 细菌Lamassu防御系统的工作模式图
中国科学技术大学博士研究生李明、博士后赵晓龙,北京化工大学博士研究生赵兴雨、李栋、熊伟佳为本论文的共同第一作者,北京化工大学生命科学与技术学院冯越教授、张怡副教授与中国科学技术大学生命科学与医学部的张凯铭教授为本文的共同通讯作者。
论文链接:https://www.nature.com/articles/s41589-025-02102-z